The Seaweed and Integrated Multi-Trophic Aquaculture (IMTA) Research Laboratory of
Dr. Thierry Chopin
S… S… S…
Ah ha, I got your attention! You thought it was Sea, Sex and Sun…! It’s even better… Seaweeds and Sex in the Sea! We even could have S… S… S… S… Seaweeds Salty Sex Stories!
Marah Hardt – researcher, writer and creative consultant from California – contacted me to talk, most respectably, about Integrated Multi-Trophic Aquaculture (IMTA). Having Googled her on the Internet, in a totally legal manner, I discovered that she also has a book in progress, Sex in the Sea, and a blog www.sexinthesea.org.
As usual, it is all about animals doing funny things in the sea! So, to break this zoological bias, I contacted Marah to propose making this entry to represent the bright side of this marine world, the photosynthetic world, and to remind the reader that seaweeds do funny things too! Yes, being photosynthetic organisms, seaweeds saw the light a long time ago and have tried all kinds of "strange" things. It is interesting to note that some are not excited by red light, but prefer the blue light district of the photosynthetically active radiation (PAR) spectrum to be stimulated!
What are seaweeds? Not surprising that they do all kinds of things!
In fact, the term “seaweeds” does not mean much! Giving a simple definition of what seaweeds are is not that simple because this heterogeneous group is only a fraction of an even less natural assemblage, the "algae". Since the time of the Greek and the Roman Empires, algae – as a misunderstood, unappreciated and underused group of organisms – have been lumped together in a very artificial manner. Algae are not a closely related group but a diverse group of organisms sharing only a few characteristics: they are photosynthesizing and provide oxygen; they do not make flowers and their anatomy is relatively simple (no roots, stems, leaves or vascular tissues, and simple reproductive structures). We now realize, especially with the progress in molecular techniques, that this “mixed bag” is completely unnatural, with no real cohesion and with species spread over most kingdoms of organisms, encompassing microscopic microalgae (like the unicellular phytoplanktonic forms) and macroscopic macroalgae (like the giant kelps that grow taller than trees) that colonize the oceans, freshwater streams, trees (associated with fungi in lichens), stones, high altitude snow in glaciers, geothermal sources and even deserts.
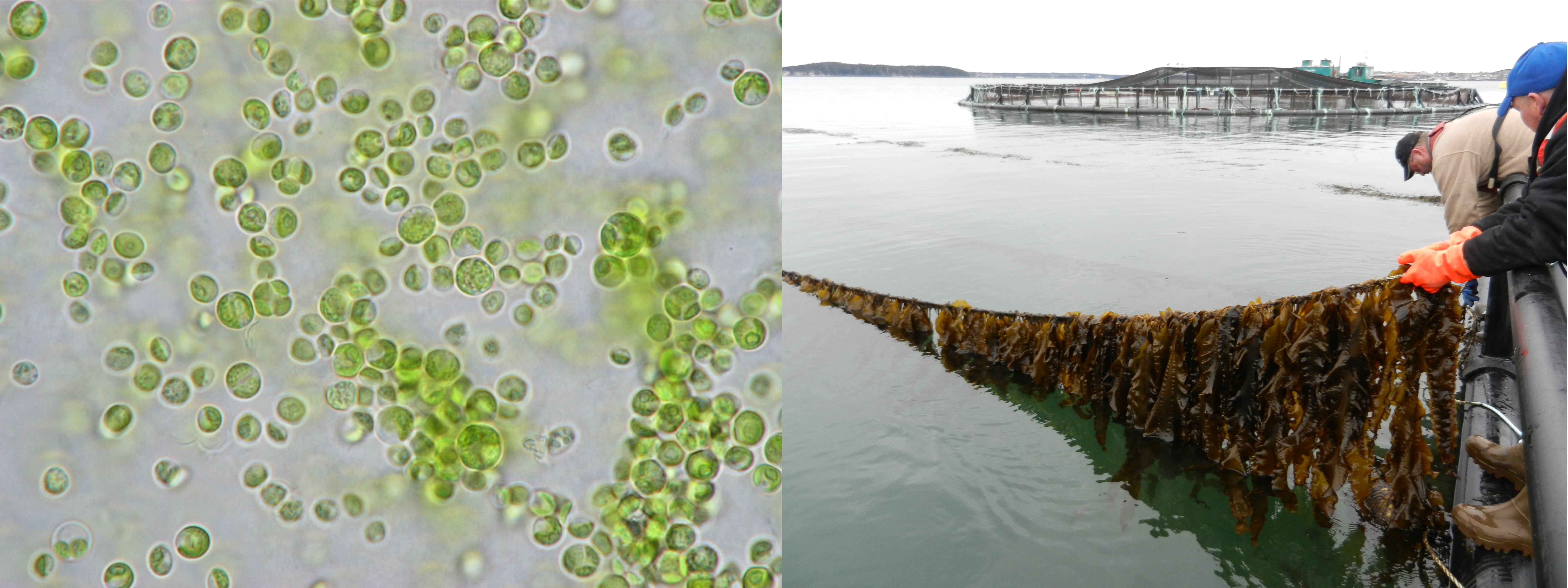
-
Phytoplankton (as seen under the microscope, on the left) or seaweeds (the kelps, on the right, grown on ropes in proximity to salmon cages at IMTA sites in the Bay of Fundy, New Brunswick, Canada) are all algae that provide key services to the ecosystem (absorbing nutrients, releasing oxygen, sequestering carbon dioxide, etc.) and can also be used in many applications in our everyday life (photo credit: Thierry Chopin).
At the molecular and ultra–structural levels, green seaweeds are closer to trees on land than they are to brown seaweeds, which are closer to some fungi… so, it is not surprising that, collectively, seaweeds are doing all kinds of things when it comes to sex and the reproductive strategies they have evolved over a very long geological time.
Understanding the life cycle of even some widely used species is quite recent
It took a very long time for scientists to understand the life cycle of some seaweeds, even ones that have been used for centuries. Progress in cultivation techniques made it possible.

-
On the left, harvesting of Pyropia (formerly Porphyra, and commonly known as nori) in Japan. The red blades are cultivated on nets that are harvested by a boat going under the nets (photo credit: Thierry Chopin). On the right, sushi rolls, with an outer layer of nori, on sale in a Japanese supermarket (photo credit: Thierry Chopin).
For example, sex in nori, the purplish wrap around shushi rolls, was only understood in 1949 by the respectable British academic Kathleen Mary Drew-Baker. Until her discovery, the cultivation of the red seaweed, Porphyra, which just recently had a name change to Pyropia, was highly empirical. She understood that the red blades, made in sheets to roll around rice and raw fish, were in fact from the sexual part of the life cycle.
Some blades were males and some were females. Because they release gametes, we call them gametophytes, male gametophytes and female gametophytes. If you are following me so far… that’s just like animals! Male gametes + female gametes = zygotes. They are the babies… but the babies do not resemble their parents… they become tiny little filaments, neither male nor female, so we enter the asexual part of the life cycle, which is, nevertheless, still very interesting and necessary! These microscopic filaments, which would be very easy prey for grazing invertebrates on the shore, are very smart! They take refuge by boring into the shells of dead shellfish, forming a reddish layer under the inner pearlescent layer of, for example, oyster shells. These filaments have been known for centuries, but were thought to be a completely different species, called Conchocelis. By cultivating this phase, Kathleen Mary Drew-Baker realised that theses filaments released spores, so Conchocelis was in fact the sporophyte of Pyropia.Suspended dead oyster shells starting to be colonized by reddish/purplish micro-filaments of the Conchocelis phase, the sporophytes, of Pyropia (photo credit: Thierry Chopin).
These spores are always released in groups of four, so they are called tetraspores; two will become male gametophytes and two female gametophytes and we are back to the sexual part of the life cycle! This discovery, by Kathleen Mary Drew-Baker, led to a major breakthrough in the commercial cultivation of nori, which, today, is worth at least US$1.34 billion.A reproductively mature blade of Pyropia purpurea from the Bay of Fundy, New Brunswick, Canada: a longitudinal line, the “zipper”, separates the reddish/purplish female part (on the left) from the pale male part (on the right) of the blade. As the male gametes are released, the remaining material of the cells gradually deteriorates and the male section of the blade disappears (photo credit: Thierry Chopin).
The number of Pyropia species is very large worldwide: I believe we were at 13 species on the east coast of Canada and New England in the USA, but molecular biologists and their DNA sequencers are always finding new ones! We have one species, Pyropia purpurea, which does things in an even more surprising way: when it is reproductive, one side has its margins nicely reddish because it keeps the female eggs in place, while the other side has whitish margins after it has shed its male reproductive organs (spermatia, because they have no flagella) to go fertilize other female parts of other individuals. The blade of Pyropia purpurea is, hence, separated by a visible line between the male and female parts. We called that line the "zipper" and it goes down as the spermatia are released and the male tissues deteriorate.
Size does not matter!
From the example above, you could conclude that the sporophytes (filaments) are the microscopic stages, while the gametophytes (blades) are the macroscopic stages of a complex life cycle. Well, in the case of Pyropia, yes, that is true; however, if I talk about the brown seaweeds called kelps, it is exactly the opposite: the large organisms are the sporophytes and gametophytes are tiny little filaments only visible under a microscope! The large brown seaweeds you can see at very good low tides are the sporophytes because they release spores (again, an asexual step, but there are tricks to make these releases abundant and successful!). Spores will germinate into small filaments (male gametophytes releasing sperm with flagella) and larger filaments (female gametophytes retaining the eggs). Sperm + eggs = zygotes… so, we are back to something you know! These zygotes develop into small little sporophytes, which become the several meter long kelps you can see at low tides, within 5-7 months.

-
The different steps in the cultivation of the kelp Saccharina latissima in the Bay of Fundy, New Brunswick, Canada. a) Collection of reproductively mature (visible dark sori in the middle of the blade) sporophytes. b) Drying sori excised from vegetative blades. c) Sori releasing spores in beakers containing seawater. d) Spore solution. e) The spore solution is inoculated on twine spooled on PVC pipes in a culture tank. f) Spores germinate into female and slender male microscopic filamentous gametophytes on the spools. g) After male gametes have fertilized female gametophytes, each zygote develops into a microscopic sporophyte. h) Spools on PVC pipes covered with microscopic brown sporophytes. i) Young sporophytes, 0.5 to 1 mm in length, ready for transfer at the aquaculture sites. j) Triple line culture system, installed in November, between compensator buoys with ropes around which the twines, “seeded” with young sporophytes, have been unspooled. k) Ropes of adult sporophytes six months later (May). l) and m) Harvesting of kelps in July (photo credit: Thierry Chopin).
This happens in nature, with the microscopic stages unnoticed at the surface of rocks; what we do when cultivating kelps is concentrate the spores in tanks and have them settle on twine instead of dispersing in the ocean, thereby increasing the chance that the sexual part of the life cycle takes place.
Now, some seaweeds seem to go through a lot of trouble to produce gametophytes and sporophytes of very different morphologies, but with no syngamy (that pretty much means no sex!) and no meiosis. They are said to have a heteromorphic asexual life cycle.
Macroscopic sporophytes of Chordaria flagelliformis in the Bay of Fundy, New Brunswick, Canada (photo credit: Thierry Chopin).
For example, in the case of the arctic to cold-temperate northern hemisphere species Chordaria flagelliformis, microscopic gametophytes occur at low temperatures combined with short days and macroscopic sporophytes occur at higher temperatures combined with long days. Unfused gametes can develop directly into gametophytes and sporophytes are able to produce parthenogenetic sporophytes or have sporophytes that repeat themselves via mitospores. So, what are the purposes of all these complicated morphological changes: better over-wintering of one form, better resistance to grazing, better…? There must be an advantage somewhere or the species would not continue in this manner. Maybe, based on the species name, better flagellation, if you are into that?!
What do you do when neither the male nor the female have flagella?
It is well known that these little flagella can be quite important for successful sex! Either they allow you to swim towards your partner or, once close, they help you to grab your partner to keep it in position. Well, in the red algae, neither the male gametes nor the female gametes have flagella… not very good for your chances of success when you have external sex in an environment that dilutes everything: seawater, waves, currents…!
Some red seaweeds are smart: they created a third generation, the carposporophyte, which acts to multiply the chances of success. OK, I think I lost you there: gametophytes, carposporophytes, tetrasporophytes… that is much kinkier than any animals could think of! Let’s take our time and breathe! Male gametophytes release spermatia carried by the current. If they are lucky they will fall on a papilla, a little protuberance on the females of the more primitive red algal species, or on an elongated filament, the trichogyne, on the females of the more advanced red algal species. Fecundation will occur and the zygote will not develop into one new individual, but will be burgeoning into a serious bump on the female gametophyte, the carposporophyte.

-
On the left, a female gametophyte of the red seaweed Chondrus crispus (commonly known as Irish moss) from the Bay of Fundy, New Brunswick, Canada. Female organs in the tissues have already been fecundated by male organs (spermatia, with no flagella, released in the water). The zygotes have been burgeoning into bumps on the female gametophytes, the carposporophytes (photo credit: Thierry Chopin). On the right, a tetrasporophyte of the red seaweed Chondrus crispus from the Bay of Fundy, New Brunswick, Canada. The dark spots are sori containing many spores in groups of four, hence the name tetraspore and tetrasporophyte. The pale spots are sori which have already released their tetraspores (photo credit: Thierry Chopin).
The carposporophyte will become a multicellular outgrowth and each of its cells will be able to release carpospores, each germinating into a tetrasporophyte. Hence, the carposporophyte simply multiplies the number of sporophytes resulting from one lucky fecundation. A multitude of carpospores will germinate into a multitude of tetrasporophytes, themselves releasing an even higher number of tetraspores germinating in a multitude of male and female gametophytes. And the circle is completed. Are you still there?!
You thought it was complicated enough; we have even better for you!
The red seaweed, Palmaria palmata, commonly known as dulse on each side of the Atlantic Ocean, complicates things even further: the female gametophyte is a microscopic form less than a millimeter in diameter, while the male gametophytes and the tetrasporophytes can reach 30-50 cm in length.
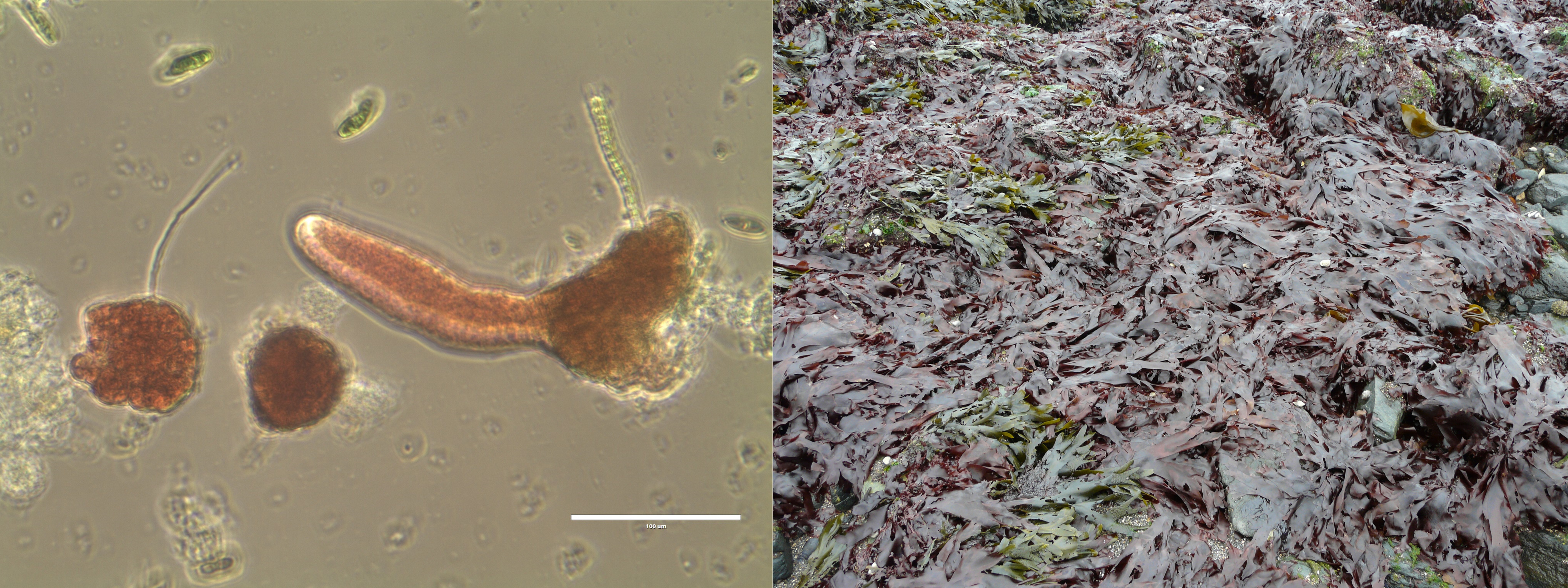
-
On the left, after 2.5 months in culture: a dwarf female gametophyte of the red seaweed Palmaria palmata (commonly known as dulse) on the left, and a male gametophyte, just starting to elongate, on the right (scale bar = 0.1 mm) (photo credit: Constanza Chianale). On the right, macroscopic blades of Palmaria palmata (i.e.male gametophytes and/or tetrasporophytes) on the shore of the Bay of Fundy, New Brunswick, Canada (photo credit: Thierry Chopin).
That makes for very “unbalanced” sex partners for a species that remains very difficult to cultivate fully to this day (we hope to change that soon). Moreover, these tiny females become sexually mature very rapidly: they produce trichogynes within a few days of existence; however, the number of trichogynes declines and they become rare on females 4-5 months old, which by then have grown into small, mis-shapen and stunted organisms. In contrast, males produce spermatia only after they are 8-12 months old.

-
On the left, male gametophytes of Palmaria palmata are also difficult to identify on the shore. When mature, they have pinkish reproductive areas and are generally slimier than tetrasporophytes (photo credit: Constanza Chianale). On the right, part of mature male gametophyte of Palmaria palmata ready to release spermatia (photo credit: Constanza Chianale).
Consequently, precociously fertile females are almost certainly unable to mate with males of the same gametangial generation and are most probably fertilized by older males from the preceding generations that have over-wintered… so, there are also Lolitas and old male perverts among seaweeds! More scientifically, it may represent an effective mechanism for maintaining heterozygosity in a population.
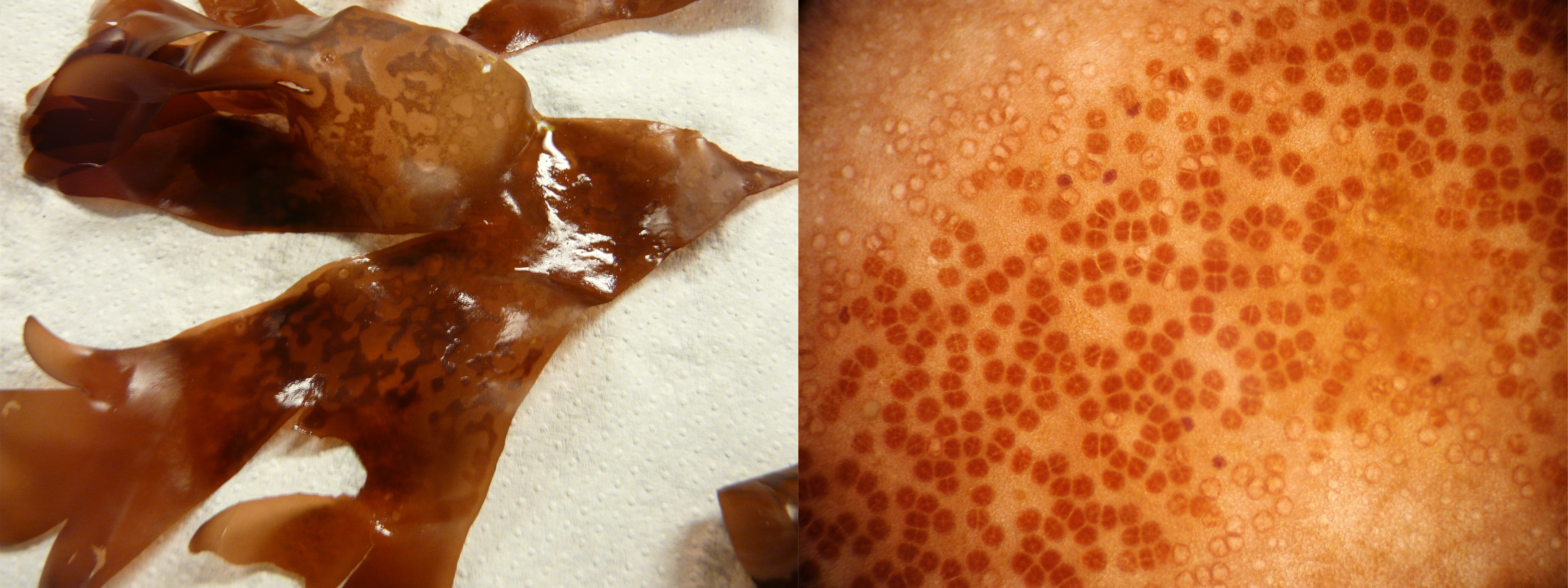
-
On the left, mature tetrasporophyte of Palmaria palmata with darker sori of tetraspores (photo credit: Thierry Chopin). On the right, details of sori of a mature tetrasporophyte of Palmaria palmata with tetraspores in groups of four (photo credit: Constanza Chianale).

-
On the left, tetraspores ready to be released from a mature tetrasporophyte of Palmaria palmata (photo credit: Constanza Chianale). On the right, jars containing solutions of tetraspores released from mature tetrasporophytes of Palmaria palmata (photo credit: Thierry Chopin).
The two short videos below show examples of an impressive spermatia release from a male gametophyte (left) and a slower release of tetraspores from a tetrasporophyte (right).
While the use of dulse as offerings by Celtic druids many centuries ago is clearly documented, the understanding of its life cycle, which is a case of extreme sexual dimorphism, is very recent: 1980!
The way it was discovered is also worth reporting: serendipity or educated intuition?! The apparent lack of females had led some scientists to doubt that P. palmata had a sexual life cycle. John van der Meer, a research scientist at the National Research Council of Canada in Halifax, Nova Scotia, Canada, decided to reopen the case and below is what he wrote to me.The sexual life cycle of Palmaria palmata (from van der Meer J.P. and Todd E.R. 1980. The life history of Palmaria palmata in culture. A new type for the Rhodophyta. Canadian Journal of Botany 58:1250-1256).
His technician, Edna Todd, and he were carefully collecting individual released spore tetrads from tetrasporophytes and culturing them in small dishes. Survival was poor, but, one day, using individuals collected on Grand Manan Island in the Bay of Fundy, they were able to get several cases with all four surviving sporelings. After a few weeks of growth, two of the spores were growing robustly and the two others were growing slowly and appeared stunted. Several months later, the two robustly growing sporelings had developed into normal-looking fertile males, whereas the two stunted sporelings barely grew. This pattern was observed for every surviving tetrad. They realized that the stunted individuals were somehow related to the missing females in the mystery; however, the puzzle was not solved yet.
Edna Todd was also culturing an interesting non-fertile green frond that had appeared spontaneously on a normal red individual. Because there was a very sharp line separating the red and green tissue, it appeared that this was most likely a mutation of a nuclear gene (mutations in chloroplast genes initially appear mottled and take more time (growth) to segregate into a unicolor sector). Edna then made what would prove to be a critical decision. To grow well, Palmaria has to be cultured in large vessels, which uses a lot of media. Mostly to save work and space, but also out of curiosity about what might happen, Edna decided to co-culture the green frond with a tetrasporophyte in a single flask. In the course of time, both became fertile, the green frond as a male plant releasing spermatia, while the tetrasporophyte was releasing tetraspores, some of which settled and grew on the bottom of the large flask. Many months later, the bottom and sides of the flask were covered with a red mat of germinating spores, holdfast discs and growing sporelings.
During the week between the Christmas and New Year holidays, Edna and John decided to do some cleaning in the lab. They decided that the Palmaria fronds should be transferred into a fresh flask and the old flask went to the sink for cleaning. After a couple of hours, John decided to retrieve it and look at the bottom and sides with a magnifying glass. To his amazement, he saw several small green discs and sporelings scattered among the many red ones. Clearly, there had been sexual reproduction involving the green male, leading to the production of a new generation of green individuals, although how it had happened was not immediately evident.
To examine the material attached to the flask more closely, and to recover some green sporelings for continued culturing, they had to break the flask, and examine fragments under a binocular microscope. Among the thousands of red discs and developing sporelings (and a few green ones) they found one bizarre-looking older individual that closely resembled the stunted ones they had obtained from culturing isolated spore tetrads, but in this case, one of its frond tips had developed into a very small, but otherwise normal-looking, fertile tetrasporophytic frond. They put this unusual individual into a large dish to collect tetraspores and in a few days it was evident that this small tetrasporophyte was the source of the green sporelings in the flask. Tetraspores from this individual yielded a 1:1 segregation of green:red sporelings, indicating that this small tetrasporophyte was the product of sexual reproduction involving the green male and the stunted individual, and was heterozygous for an inherited recessive green mutation.
They also observed that sporelings with "hyaline hairs" (which are produced by many algae during early sporeling development) and sporelings without them were also present in a roughly 1:1 ratio. This seemed somewhat odd. Upon closer inspection, it became evident that these "hairs" were actually very long trichogynes (hair-like projections) emerging from carpogonia (egg cells) on what had to be fertile females... even though they were less than a week old. These observations were shown to be correct by exposing tiny green, presumptive, females to spermatia from normal red males. Fertilization took place (spermatia became attached to the trichogynes) and shortly afterwards, small red fronds began to develop on the tiny, green, female individuals. After some further growth, the base of these small red fronds developed files of red cells that grew over the green females to the substratum to form a holdfast disc, and before long, the females disappeared completely, overgrown by the developing red tetrasporophytic fronds, which explained why they had never been found in nature.
Quite some detective work and quite a lesson! Precocious sexuality of little Palmaria females with their mother’s male siblings, who have finally reached maturity, condemns them to being buried by their offspring’s overgrowing holdfast on the same rocky spot they occupy, hence leading people to believe for a very long time that they never existed!
When related species do things differently at different heights on the shore
On both sides of the Atlantic Ocean, two related species of brown seaweeds, Fucus spiralis and Fucus vesiculosus, do things very differently just a few meters apart.
High in the intertidal zone, Fucus spiralis is a monoecious species. Another scientific word… but not so complicated if we look at its Greek roots: mono = one and oikos = house. As I explain to my students, who generally do not know much about Greek or Latin etymology, a monoecious species is like a co-ed residence at university: both males and females live under the same roof. So, a monoecious species is a species which "houses" both male and female organs on the same individuals. In F. spiralis, the male and female reproductive organs are contained within the same conceptacles. But just as co-ed residence does not mean constant bacchanals, F. spiralis has found a way of guaranteeing and maintaining biodiversity by creating mating incompatibility between male and female organs of the same conceptacle.Fucus spiralis, growing in the upper intertidal zone of the Bay of Fundy, New Brunswick, Canada. The reproductive parts, the receptacles, are visible; they contain monoecious conceptacles with both male and female reproductive organs (photo credit: Thierry Chopin).
Lower in the intertidal zone, Fucus vesiculosus is a dioecious species. This time, there are residences for males and residences for females, i.e. some individuals have conceptacles containing only male reproductive organs and other individuals have conceptacles containing only female reproductive organs. We are back to what happens with the majority of animal species which are dioecious (also called gonochoric), i.e. the state of having just one of at least two distinct sexes in any one individual organism. It is interesting to note that the majority of terrestrial plant species are, instead, bisexual (or hermaphrodite) with flowers possessing a mixture of male and females attributes (stamens and ovaries). Maybe only 6 % of terrestrial plants are fully dioecious.Fucus vesiculosus, growing in the middle of the intertidal zone of the Bay of Fundy, New Brunswick, Canada. The vesicles are particularly abundant on this example; they are for floatation purposes and are not to be confused with reproductive structures (photo credit: Thierry Chopin).
The receptacles of Fucus are the big swollen parts, containing many conceptacles, at the ends of individuals, and a treat that children walking on the shore enjoy popping. Without realizing it, they have provided a very simple method for determining the sex in Fucus vesiculosus (and other dioecious species of Fucus), without resorting to delicate dissections. Just pop a receptacle: if you get a greenish gel substance between your fingers, you have a male; if you get an orangeish gel substance, you have a female (the eggs are much bigger than the sperm cells and, as in many species, they are the ones providing the reserves, including the brown pigments in this case).
Why do two species belonging to the same genus, Fucus, and separated vertically on the shore by only a few meters, do things so very differently? That is an enigma: they have chosen different reproductive strategies but both are very successful in their own niche.
Like many marine organisms, seaweeds invest massively in the production of reproductive material
On calm days in the spring, the green seaweed Ulva lactuca (commonly known as sea lettuce) can release thousands and thousands of gametes or spores in the Bay of Fundy, New Brunswick, Canada, which create this foamy green juice in the seawater. The margins of the blades become whitish as they release either anisogamous gametes (the female gametes are bigger than the male gametes, but otherwise similar) or spores (the gametophytes and the sporophytes look alike) (photo credit: Thierry Chopin).
Does that mean that they "think" only about sex, or even asexual encounters?! One has to consider that mostly external sexual acts in an environment that disperses things as soon as they are released, are not conducive to a high chance of reproductive success. The strategy of many marine organisms, whether animals or algae, is, therefore, to invest a lot of energy in the production of reproductive material with the hope that there will be a few successful fecundations or spore germinations.
Let's take, for example, the case of Ascophyllum nodosum, commonly known as rockweed or bladderwrack. This is a very successful colonizer in the intertidal zone, in mostly sheltered to semi-exposed habitats. Within 3 weeks, around the month of June, it will release all its reproductive organs from receptacles that have become very swollen.

-
On the left, in the Bay of Fundy, New Brunswick, Canada, the intertidal zone is mostly covered by a very successful colonizer, the brown seaweed Ascophyllum nodosum (commonly known as rockweed) (photo credit: Thierry Chopin). On the right, in June, the receptacles of Ascophyllum nodosum in the Bay of Fundy, New Brunswick, Canada, are very swollen and ready to release the gametes, contained in the conceptacles, over a period of a few weeks. During that period, walking on the shore is more slippery than usual (photo credit: Thierry Chopin).
These receptacles will even be shed, representing in some cases as much as 25 % of the weight of the individuals being detached. Spending 25 % of its weight on reproductive structures is itself quite remarkable, but, from an ecological point of view, this also represents a significant spike of organic matter in the coastal environment within a few weeks. These tons and tons of organic matter will decay and return to inorganic forms, which represent nutrients, i.e. fertilizers, which are important for the proper functioning of the ecosystem. In fact, where rockweed is harvested, like on the east coast of Canada, the harvesting season starts after this huge reproduction and fertilization event has taken place. The industry is not very interested in harvesting individuals with ripe receptacles: they are full of water, so why pay for water when you dry the material later? Moreover, they are not only watery, but also full of sugars that make for a gluey material clogging the machineries. Yuk, yuk, yuk!
Another example of copious release of reproductive material in the sea is what happens with kelps. We are cultivating two species of these large brown seaweeds, Saccharina latissima and Alaria esculenta, as the inorganic extractive component of Integrated Multi-Trophic Aquaculture systems (to know more about IMTA, please, peruse my website at www2.unb.ca/chopinlab/).

-
On the left, a line of kelp (Saccharina latissima) cultivated near an Atlantic salmon (Salmo salar) cage at an integrated multi-trophic aquaculture (IMTA) site of Cooke Aquaculture Inc. in the Bay of Fundy, New Brunswick, Canada (photo credit: Thierry Chopin). On the right, a line of kelp (Alaria esculenta) cultivated at an integrated multi-trophic aquaculture (IMTA) site of Cooke Aquaculture Inc. in the Bay of Fundy, New Brunswick, Canada (photo credit: Thierry Chopin).
We have calculated that 100 individuals of S. latissima can release as many as 8.8 billion spores, and 100 individuals of A. esculenta as many as 7.2 billion spores, after we cajole them overnight and play with temperature and moisture to optimize these major outbursts of spores.
Containers with spore solutions of the kelps Alaria esculenta (left) and Saccharina latissima (right) ready to be inoculated on twines (photo credit: Thierry Chopin).
While the answer to what could be not very successful external fertilization seems a massive orgy of released reproductive organs, some seaweeds have also “learned” how to better their odds by releasing gametes under optimal hydrodynamic conditions for sperm-egg encounters. For example, different species of Fucus release their gametes when the water is calm, on sunny days and when the concentration in dissolved inorganic carbon (DIC) around them is low. Sunny days mean high levels of photosynthesis and concomitant fixation of carbon, leading to low levels of DIC in the thin boundary layer surrounding the Fucus blades, which remains unstirred (i.e. not replenished) by still water. Talk about coordinated signals converging towards optimized stimulating conditions!
And you thought that sex in seaweeds would be boring…
Well, hopefully these few examples showed you that is not the case! Seaweeds, members of this vast and unnatural grouping that constitutes the algae, remain organisms that are low in the evolution tree. Consequently, it is not surprising that they have tried all kinds of things that jostle our mind so biased toward animalistic schemes.
We have to consider that they are all different trial strategies to increase reproductive success and colonization in an environment that can be quite hostile. Moreover, all these different species may be at different stages in their evolution and it is difficult for us humans, only present on this planet for a short time, to appreciate whether these trials will lead to successful adaptations or odd dead ends.
So, next time you think of Sex in the Sea, remember that seaweeds have been much more "imaginative" than animals, with their "boring" diplontic life cycle!
The pictures on this page, and throughout the entire website, are property of their owners. As such, they cannot be used, or reproduced, without their expressed consent.
Copyright © 2006
UNB |  Accessible Version | Site Map
Accessible Version | Site Map